|
Pinguicula (Lentibulariaceae) : les espèces de climat froid de l'hémisphère nord, morphologie, biologie et culture.
D'après une présentation du Dr Juerg F. STEIGER, 2nde conférence de l'ICPS, Bonn, Juin 1998,
| ||||||||||||||||||||||||||||
|
Pinguicula balcanica Vardoussia, centre Grèce.
|
|
|
(Photos Jurg F Steiger)
|
|
|
Pinguicula grandiflora f. pallida, Col de la Faucille, Jura, France. |
Pinguicula nevadensis, Mont Veleta, Sierra Nevada, Espagne. |



INTRODUCTION :
L'origine géographique du genre Pinguicula n'est pas connue car on ne dispose pas de fossiles de ces plantes trop fragiles. Par contre, quelques éléments permettent de supposer que ce genre était déjà représenté il y a une cinquantaine de millions d'années (soit au Miocène). A cette époque, le continent américain et le continent eurasien n'étaient pas encore totalement séparés. Quelques passerelles terrestres dans le Nord atlantique et les îles autorisaient encore les migrations intercontinentales de plantes. On admet à ce jour l'existence de deux centres de distribution qui sont à l'origine des zones de distribution actuelles : un se trouve autour de la Mer Méditerranée, l'autre autour des Caraïbes.
Quelques populations subsistent en Eurasie, dans les régions du nord de la zone méditerranéenne, d'autres ont migré vers le nord (y compris jusqu'en Islande) ainsi que vers l'est pour atteindre le Japon et l'Himalaya. En Amérique, quelques populations sont situées à l'intérieur mais également autour de la zone des Caraïbes, d'autres ont migré vers le nord et le nord-ouest, d'autres enfin, vers le sud en suivant la cordillère des Andes pour arriver jusqu'à la terre de feu.
Le genre Pinguicula n'est pas représenté en Afrique (excepté dans le nord du Maroc), en Indonésie, en Australie et en Nouvelle-Zélande.
Lors de changements de conditions environnementales, trois stratégies d'évolution sont observées chez les plantes : migration, adaptation ou extinction. Pendant plusieurs millions d'années, la plupart des zones terrestres vivaient sous un climat subtropical, mais il y a environ un million d'années, une succession de périodes glaciaires a obligé des centaines d'espèces de plantes à migrer et/ou à s'adapter. Il en a résulté une redistribution complète des zones de distribution et des génotypes. Des études géographiques, morphologiques et la réalisation des caryotypes de Pinguicula devraient apporter des éléments sur les différents types de stratégies qui ont été adoptées à l'intérieur du genre (adaptation, migration et différenciation).
Dans les années 30, HAGERUP, TISCHLER et d'autres auteurs sont arrivés à l'hypothèse suivante (loi HAGERUP-TICHLER) : des périodes environnementales difficiles, particulièrement instables, imprévisibles, feront augmenter le nombre de chromosomes chez les plantes (c'est le cas par exemple d'une alternance de périodes glaciaire et interglaciaire, périodes de sécheresse, des amplitudes extrêmes de températures, d'intenses radiations UV). Cela signifierait que des plantes avec un nombre élevé de chromosomes seraient plus aptes à supporter une ou plusieurs périodes glaciaires, alors qu'au contraire un petit nombre de chromosomes prouverait le besoin pour ces plantes d'avoir des conditions environnementales relativement stables (selon HAGERUP, 1932).
| Environnement stable | Environnement instable | |
| Nombre de chromosomes FAIBLE (gamètes haploïdes). | Possibilités de recombinaisons suffisantes. | Possibilités insuffisantes de recombinaisons --> survie génétique à long terme non garantie. |
| Nombre de chromosomes ÉLEVÉ (gamètes diploïdes) | Pas de nécessité d'importantes possibilités de recombinaisons. | Besoin d'importantes possibilités de recombinaisons. |
Ainsi, si l'on suit ce principe, on peut considérer que des conditions environnementales difficiles et imprévisibles entraîneront une augmentation du nombre de mutants. Mais une mutation défavorable dans un des gamètes, pourrait causer un handicap sélectif chez les descendants, particulièrement si elle affecte un gène dominant.
Habituellement, les grains de pollen et les gamètes femelles sont haploïdes, ce qui donne lors de la fécondation une cellule diploïde.
De plus, on a découvert que pour d'autres familles de plantes, il y a en moyenne un pour mille spécimen tétraploïde à cause d'un mauvais déroulement de la phase de méiose chez des gamètes parentales ou durant la phase de fusion des deux gamètes (BRIGGS et WALTER 1973). Si une telle mutation s'opère dans une gamète diploïde, il est peu vraisemblable que cette même mutation affecte également les gènes correspondants dans les autres chromosomes homologues. Lors de la fusion des deux gamètes diploïdes, formant alors une cellule tétraploïde, cette mutation sera présente mais uniquement chez un des quatre chromosomes homologues. Ainsi, pour des plantes polyploïdes, les trois chromosomes sains protégeront la plante de mutations à son désavantage. De plus la représentation multiple du code génétique pour un même caractère engendrera une meilleure tolérance écologique grâce à l'hétérogénéité des gènes homologues.
Les pinguiculas peuvent être regroupées suivant leur type de croissance en quatre groupes : tropical/homophylle, tropical/hétérophylle, tempéré/homophylle et tempéré/hétérophylle. Le groupe du type tropical forme des feuilles carnivores toute l'année et celui du type tempéré passe l'hiver sous la forme d'un hibernacle. Dans le type homophylle, seul un type de rosette est formée : toutes les feuilles de la rosette ont morphologiquement les mêmes caractéristiques. Le type hétérophylle par contre, forme deux rosettes différentes : soit une plus grande rosette en été et une plus petite, morphologiquement différente en hiver (type tropical/hétérophylle), soit une plus petite rosette au printemps suivi par une rosette plus grande en été avec une morphologie différente des feuilles, qui plus tard formeront l'hibernacle (type tempéré/hétérophylle).
|
Tropical |
Tempéré |
|
|
Homophylle Un seul type de rosette |
P. jackii | P. vulgaris |
|
Hétérophylle Deux types différents de rosettes |
P. gypsicola | P. vallisneriifolia |
La présentation concernera uniquement les espèces de Pinguicula des régions tempérées, (sub)alpines et (sub)arctiques de l'Eurasie mais sera complétée de trois espèces présentes au nord-est et au nord de L'Amérique. Les espèces présentées appartiennent aux sous-genres Pinguicula,Temnoceras et Isoloba.
A. Sous-genre Pinguicula
Il comprend (par ordre alphabétique) les espèces suivantes : P. balcanica,corsica , dertosensis (submediterranea), fiorii, grandiflora, leptoceras,longifolia, macroceras , mundi, nevadensis, vallisneriifolia, villosa et vulgaris.
Toutes les espèces de ce groupe hibernent sous la forme d'un bourgeon hivernal (hibernacle) qui forme à sa base des petits bourgeons (gemmes) permettant la reproduction végétative. Leurs feuilles et racines disparaissent en hiver. Le nombre de chromosomes de ces espèces est à chaque fois un multiple de 8.
1. Pinguicula corsica Bern. & Gren. Ex Gren. & Godr. 1850

Cette espèce bien distincte est endémique de la Corse et n'a jamais été trouvée ailleurs. Il y a six millions d'années, lorsque le niveau de la mer Méditerranée était inférieur à celui qu'elle occupe de nos jours, la Corse n'était pas encore séparée du continent par les eaux de l'Atlantique entrées par le détroit de Gibraltar (Illustr. Wiss. 1998). Il est vraisemblable qu'une ou plusieurs anciennes populations de Pinguiculaexistaient dans les zones montagneuses du sud de l'Europe. Quand la mer Méditerranée a finalement séparé la Corse et la Sardaigne du continent, elle a aussi isolé les pinguiculas de Corse. La Corse est alors restée plus ou moins épargnée par plusieurs périodes glaciaires. Son climat de type insulaire était probablement assez prévisible pour les plantes. Il n'a donc pas été nécessaire à cette espèce de s'adapter à des changements fondamentaux de son environnement ou de conquérir de nouveaux territoires. Il n'y avait donc ainsi aucune raison obligeant cette espèce à accroître son nombre de chromosomes. Avec 2n=16, P. corsica est considérée comme étant une des plus vieilles espèces de tout le sous-genre.
P. corsica est caractérisée par des feuilles plutôt larges et grandes (plus de 40 mm de long et plus de 25 mm de large), par la gorge étroite de la fleur, un éperon mince et par les pointes étroites du calice mais qui restent bien séparées. Les fleurs peuvent varier en couleur depuis un pratiquement blanc jusqu'au bleu-pâle. La taille de la fleur est de 25mm.Pinguicula corsica pousse dans des prairies humides alpines et sub-alpines, autour de petits torrents ou de lacs, au niveau et au dessus de la limite boisée, descendant parfois dans des gorges fraîches et souvent entre 1000 à 2400m d'altitude. Les sites les plus élevés sont recouverts par la neige pendant 4 à 7 mois. Le substrat de tous les sites est composé de granite, mais en culture, P. corsica peut aussi pousser sur un substrat calcaire. En fait, on peut dire qu'elle ne pousse pas sur un sol siliceux par préférence après un long processus de sélection naturelle, mais seulement parce qu'aucun autre substrat n'était disponible sur l'île.
2. Pinguicula nevadensis (Lindberg) Casper 1962
Il s'agit encore d'une espèce bien distincte, endémique des zones alpines de la Sierra Nevada dans le sud de l'Espagne. Ses feuilles sont aussi relativement larges, de forme similaire à celles de P. corsica mais plus petites (25 mm de long, 20mm de large) ; les fleurs sont relativement petites (13mm), variant du violet pâle au blanchâtre, l'éperon est plus fin et plus courbé que chez P. corsica. Cette espèce se rencontre bien au dessus de la limite forestière et pousse sur de nombreux sites près de petits torrents et de lacs alpins sur du substrat granitique entre 2160m et 3100m d'altitude. Bien que P. nevadensis soit exposé à de très intenses radiations solaires, ses feuilles n'ont qu'une très légère pigmentation rougeâtre. Les sites les plus élevés sont recouverts par la neige durant 5 à 7 mois. Une étude des proies qu'elle capture a été publiée par Zamora (Zamora, 1990).
Pendant les précédentes périodes glaciaires et périodes interglaciaires, les chaînes de montagnes du sud de l'Espagne n'ont pas été exposées à des changements de climat aussi brusques que dans les Alpes ou dans la plupart des zones du centre et du nord de l'Europe. Ainsi, le faible nombre de chromosomes de P. nevadensis (2n=16) n'est pas une surprise et indiquerait que nous nous trouvons en présence d'une vieille espèce qui n'a pas été obligée de migrer pour conquérir de nouveaux territoires.
3. Pinguicula leptoceras Reichenbach. 1823
P. leptoceras se rencontre dans des sites humides subalpins et alpins dans les Alpes à l'ouest de l'Autriche, au nord et nord-ouest de l'Italie, en Suisse et dans le sud-est de la France. Ses feuilles varient considérablement en taille et sont parfois pigmentées dans certains sites ensoleillés. La principale caractéristique de P. leptoceras est son calice : la pointe supérieure centrale est souvent très large, celles du bas sont bien séparées en deux pointes minces formant entre elles un angle de 90o ou plus. Elle se distingue aussi de la plupart des autres pinguiculas européens par sa capsule de graines de forme globulaire. On ne sait pas clairement pourquoi Reichenbach a appelé cette espèce 'leptoceras' (=éperon mince) car l'éperon est souvent large et fin, même dans le site de l'holotype où j'ai étudié des centaines de fleurs. Les fleurs mesurent environ 22 mm de long et les lobes de la corolle se chevauchent. Dans de nombreux sites, des fleurs avec 6 à 7 lobes de corolle et pointes de calice surnuméraires sont très fréquentes. Dans ses zones de distribution à l'Est, les fleurs ont souvent une couleur bleu-violet foncée parfois avec de 1 à 3 petites taches blanches sur les lobes inférieurs de la corolle. Sur le site de l'holotype, au centre de la Suisse, la plupart des spécimens ont trois taches blanches bien distinctes, dans les Alpes Maritimes, les fleurs sont bleu-pâle avec trois grandes taches blanches. P. leptoceras se rencontre souvent entre 1800m et 3000m d'altitude et descend rarement au-dessous de 1500m. Dans les Alpes-Maritimes, elle pousse sur des roches calcaires ; dans les zones Nord et Est de sa répartition, elle semble préférer un substrat siliceux. P. leptoceras appartient au groupe des tétraploïdes (2n=32), ce qui pourrait signifier que l'on serait en présence d'une espèce jeune. On la rencontre occasionnellement en compagnie de P. alpina et P. vulgaris. Sur quelques sites où ces plantes sont communes, j'ai enregistré des hybrides naturels de P. leptoceras avec P. vulgaris mais jamais avec P. alpina.
P. leptoceras a aussi été répertoriée dans les Apennins en Italie. J'ai visité un des sites (subalpin, limite alpin, au dessus de Carrara) où l'on a enregistré P. leptoceras mais ce que j'y ai trouvé est à coup sûr niP. leptoceras ni P. longifolia ssp. reichenbachiana. En fait, la répartition deP. leptoceras est probablement limitée aux Alpes stricto sensu. Les pinguiculas des Apennins nécessitent une étude ultérieure.
4. Pinguicula balcanica Casper 1962
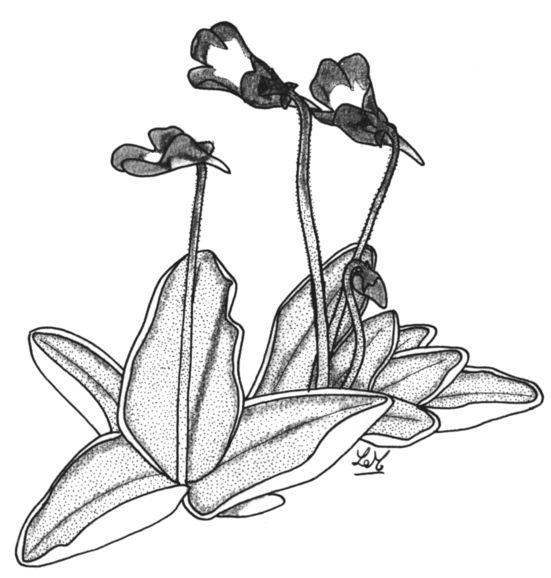
P. balcanica est à proprement parler, le complémentaire à l'Est de P. leptoceras. Il est tétraploïde (2n=32) et pousse dans des sites humides subalpins et alpins dans les Balkans, en Albanie, Macédoine et en Bulgarie jusqu'à l'extrême sud des Pîndes en Grèce (en 1995 Jan Schlauer et moi-même trouvâmes dans la chaîne Vardoussia au centre de la Grèce, le site le plus au sud répertorié à ce jour). En Bulgarie, la limite de distribution nordique a besoin d'être définie plus précisément (Peev, 1997). Un de ses traits caractéristiques est un chevauchement des lobes supérieurs de la corolle qui sont très proches les uns des autres.
La coloration de la corolle est presque similaire à celle de P. leptoceras mais les taches blanches sont moins visibles. La capsule de graines est de forme conique en pointe. P. balcanica pousse entre 1700 et 2400m, c'est à dire une altitude bien supérieure à la limite forestière, aussi bien sur substrat granitique que sur substrat calcaire. Casper décrit cette espèce comme étant hétérophylle ce que je peux difficilement confirmer. Quoiqu'il en soit, les feuilles des spécimens que j'ai collecté dans les monts Vitosha (sud de Sophia) sont si différentes de ceux des deux sites de Grèce qu'il n'est pas invraisemblable que les Balkans puissent abriter deux sous-espèces ou même espèces.
Casper décrit également une espèce pétrophylle, P. balcanica var. tenuilaciniata de Bosnie, Hercégovie et Monténégro avec des feuilles similaires à celles de P. longifolia (Casper, 1962). Il est vraisemblable que cette variété soit hétérophylle. Je n'ai jamais vu cette plante qui pourrait être proche ou identique, d'une nouvelle Pinguicula récemment découverte par Poldini dans le Nord-est de l'Italie.
5. Pinguicula fiorii Tammaro & Pace 1987

Cette espèce découverte en 1987, est endémique des montagnes Maiella dans le centre de l'Italie (Tammaro & Pace, 1987). Trois sites sont enregistrés à ce jour : les deux plus bas dans la zone forestière et un dans la zone alpine. Il s'agit d'une remarquable distribution entre 750 m et 2470 m. Tous les sites sont sur substrat calcaire. P. fiorii est bien distincte d'espèces proches, avec lesquelles elle diffère par plusieurs points : par son calice plus grand et imposant, par l'absence de toute tache blanche sur les lobes inférieurs de la corolle. Les lobes de la corolle sont violet soutenu, parfois presque bleus.
Contrairement à une autre espèce connue en Italie (P. longifolia ssp. reichenbachiana), cette espèce est homophylle. Morphologiquement, il n'y a aucune relation aussi bien entre P. corsica et balcanica qu'avec la famille des P. longifolia.
En dépit d'une description et illustration relativement succinctes de cette découverte, il s'agit certainement d'une nouvelle espèce. Le nombre de chromosomes indiqué est de 2n=32 mais basé sur un protocole d'étude insatisfaisant.
6. Pinguicula grandiflora Lamarck 1789

P. grandiflora a plusieurs aires de répartition bien distinctes depuis le sud-ouest de l'Irlande au plateau central de la France, les versants français et espagnol des Pyrénées et quelques sites de la côte atlantique du nord de l'Espagne. Une telle grille de répartition, résultat de migrations pré-, inter- et post-glaciaires sont assez typiques de plantes qui ont des besoins pour leur croissance très spécifiques. Webb pense que P. grandiflora a survécu à la dernière période glaciaire dans le sud-ouest de l'Irlande, qui a été probablement préservé des glaces pendant toute cette période (Webb, 1952). Comme on pouvait s'y attendre avec une telle répartition, P. grandifloran'est pas diploïde mais tétraploïde (2n=32). Il s'agit d'une espèce bien distincte caractérisée par une relative vigueur et de grandes fleurs (plus de 35-45mm) avec des veines bien caractéristiques. Les bords des lobes de la corolle sont ondulés. P. grandiflora demande un taux d'humidité important et préfère pousser dans des sites moussus inondés par de l'eau froide tout l'été. Il ne pousse pas dans de l'eau stagnante ou dans des sites où le substrat chauffe trop en journée.
La forme 'normale' violette est distribuée dans tous les sites mentionnés ci-dessus excepté en Savoie. En Irlande, elle atteint pratiquement le niveau de la mer ; dans le Jura et les Pyrénées, elle atteint une altitude de 1600m et un peu plus. Elle pousse habituellement dans un substrat terreux, calcaire ou sur des falaises calcaires suintantes ; exceptionnellement, on la trouve aussi sur substrat de granite dans quelques sites des Pyrénées. Dans des sites ensoleillés, la bordure des feuilles, la tige et le calice peuvent présenter une coloration foncée.
Concernant l'existence tant décriée de P. grandiflora au Maroc, Casper (communication personnelle en 1998) croit que ce spécimen a été collecté ailleurs et étiqueté par erreur comme étant originaire d'un site marocain (voir aussi Schlauer 1994)
6b Pinguicula grandiflora f. pallida (Gaudin) Casper 1962
La forme violet pâle, dite forme pallida, est marquée aussi par l'absence de pigmentation foncée de la tige et du calice comme chez la forme normale. Elle pousse occasionnellement parmi des spécimens de coloration foncée dans quelques sites du Jura de 1000m à 1500m d'altitude. Elle représente à peu près 20% des spécimens. Il y a aussi un site où l'on ne rencontre que des spécimens de forme pallida. Cette forme pâle est vraisemblablement due à une mutation (mais n'est pas un albinos stricto sensu) qui a une bonne potentialité d'occurrence puisqu'elle est régulièrement enregistrée depuis 1928. Les spécimens pâles ont aussi un éperon plus fin, donnant l'impression que cette altération génétique qui mène à la mutation de couleur serait polyphénotypique.
Une forme entièrement blanche a été trouvée dans les montagnes Burren au sud de Galway Bay (Co. Clare) en Irlande par Webb en 1956 et plus tard par Nelson en 1991, qui l'a décrite sous le nom de P. grandiflora f. chionopetra(Nelson 1993). J'ai vu un spécimen vivant de là-bas qui n'était pas pâle comme f. pallida mais bien blanc. Des P. grandiflora blanchâtres ou très pâles ont aussi été trouvées par d'autres auteurs dans le Co. Kerry, mais la forme blanc pure semble être très rare. Il y a plusieurs points de vue suivant qu'on les considère comme des albinos occasionnels comme ils s'en rencontre parmi des espèces de coloration violettes, ou s'ils doivent rejoindre le terme de forme.
6c. Pinguicula grandiflora ssp. rosea (Mutel) Casper 1962
Pinguicula grandiflora ssp. rosea est endémique des montagnes calcaires autour de Grenoble (France). Dans cette zone, la forme normale n'est pas présente. Il a toutes les caractéristiques de P. grandiflora , mais la fleur est plus petite, l'éperon est plus court et pourpre, et la couleur de la corolle est rose pâle avec des traits marqués sur la gorge. Les bords de la corolle sont souvent beaucoup plus ondulés que chez la forme normale.
A SUIVRE...
dans DIONÉE n°48
DIONÉE 47 - 2002
![]()